Адаптация штаммов-продуцентов к стабильным изотопам.Статьи / Полупрепаративный микробиологический синтез биологически активных соединений, меченных стабильными изотопами водорода и углерода / Адаптация штаммов-продуцентов к стабильным изотопам.Страница 4
'Данные (1-10) приведены для В. methylicum, не адаптированного к средам с высоким содержанием дейтерия.
Данные 10' приведены для адаптированного В. methylicum.
Выращивание проводили в периодических условиях в колбах Эрленмейера объемом 500 мл с наполнением средой до 100 мл при 35-370С в условиях интенсивной аэрации на орбитальном шейкере. 2Н -и 13С-меченые аминокислоты выделяли в чистом виде фракционированием культуральных жидкостей, включая диализ через полупроницаемую коллодиевую мембрану, обработку метанолом (-50С), ионнообменную хроматографию на катионообменнике UR-30 (элюент 0.2 н натрийцитратный буфер), химическую дериватизацию в метиловые эфиры Днс- и Кбз-производных аминокислот, разделение методом ОФ ВЭЖХ на октадецилсилановой смоле Сепарон С18 (элюент ТФУ-ацетонитрил, 80:20).
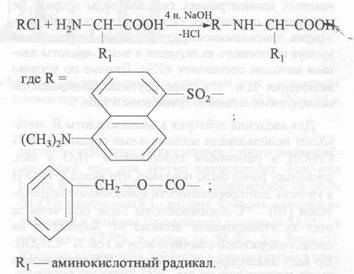
Летучие производные аминокислот синтезировали первичной химической обработкой культуральной жидкости Днс-хлоридом (Кбз-хлоридом) в 4 н растворе NaOH в ацетоне, используя пяти-кратный избыток реагента. Для лизина, гистидина, тирозина, серина, треонина и цистеина наряду с моно-производными синтезировались ди-Кбз,(Днс)-лизин, ди-Кбз,(Днс)-гистидин, О,N-ди-Кбз,(Днс)-тирозин, O,N-ди-Кбз,(Днс)-серин, O,N-ди-Кбз,(Днс)-треонин и N,S-ди-Кбз,(Днс)-цистеин. Из аргинина синтезировался три-Кбз,(Днс)-аргинин. Метиловые эфиры производных аминокислот синтезировали в условиях, исключающих обратный изотопный (Н-2Н)-обмен в ароматических аминокислотах за счет вторичной дериватизации по карбоксильной группе диазометаном. Для лизина и тирозина пики [М]+. соответствовали метиловым эфирам ди-производных аминокислот - a, e-ди-Кбз, (Днс)-лизину и О, N-ди-Кбз, (Днс)-тирозину). Конечный выход 2Н -и 13С-меченых аминокислот составил 65-90, а хроматографическая чистота - 87-95%.
Во всех анализируемых образцах культуральной жидкости наряду с основными секретируемыми аминокислотами обнаружены 5-6 ммоль аланина, валина, лейцина/изолейцина и фенилаланина. В культуральной жидкости M. flagellatum в дополнение к вышеобозначенным аминокислотам также фиксировался глицин.
2
Н- и 13С-меченые суммарные белки
получали при выращивании метилотрофных бактерий в условиях аналогичным получению секретируемых аминокислот. Сбор урожая клеток проводили на стационарной фазе бактериального роста (D540 = 2.0). Содержание белка в биомассе достигало 45% при 50%-ной конверсии метилового спирта. При выделении суммарных белков биомассы учитывалось наличие примеси нуклеиновых кислот, углеводов, липидов и пигментов. Нуклеиновые кислоты выделяли фенольным методом, углеводы - осаждением EtOH из 0.14 м NaCl, липиды и пигменты - экстракцией смесью органических растворителей хлороформ-МеОН-ацетон (2:1:1). В зависимости от таксономического рода штамма и содержания посторонних примесей реализовались различные подходы их выделения, включая высаливание белка 60%-ным сульфатом натрия, солюбилизацию ДДС-Na и фракционирование органическими растворителями (метанол, ацетон), ионнообменную хроматографию на катионнообменнике UR-30 и ОФ ВЭЖХ на Силасорбе С18 (степени хроматографической чистоты 93-96 при выходах 75-89%).
2
Н-меченый БР
. Основные этапы выделения 2Н-меченого БР (выход 8-10 мг из 1 г бактериальной биомассы, активность 85-90%) заключались в наработке 2Н-меченой биомассы в синтетической среде (4.3 м NaCl) с 2Н-мечеными аминокислотами, получении фракции ПМ, отделение от низко-и высокомолекулярных примесей, клеточной РНК, каротиноидов и липидов, фракционирование солюбилизированного в 2%-ном растворе ДДС-Na белка метанолом (-50С) и очистку на Сефадексе G-200 (0.09 м Трис-боратный буфер (рН 8.35) с 0.1%-ным ДДС-Na и 2.5 мМ ЭТДА). Образование ретинальпротеинового комплекса приводило к батохромному сдвигу в спектре поглощения БР (рис. 3): основная полоса при максимуме поглощения 568 нм, вызванная световой изомеризацией хромофора по С13=С14-кратной связи определяется наличием транс-ретинального остатка ретиналя БР568, дополнительная малоинтенсивная полоса при 412 нм характеризует незначительную примесь образующейся на свету спектральной формы M412 c депротонированной альдиминной связью между остатком транс-ретиналя и белком, а полоса при 280 нм определяется поглощением ароматических аминокислот в полипептидной цепи этого белка (для чистого БR соотношение D280/D568 равно 1.5:1).
Смотрите также
Хром (Cromium), Cr
Хром встречается в природе в основном в виде хромистого железняка Fe(CrO2)2 (хромит железа). Из него получают феррохром восстановлением в электропечах коксом (углеродом):
FeO·Cr2O3 + 4C → ...
Тиолы
В современной химии
одной из актуальных проблем является получение реагентов для проведения
различных синтезов. С каждым годом повышаются требования к чистоте реагентов.
Кроме того, немалов ...
Комплексные соединения, их биологическая роль (на примере хлорофилла и гемоглобина)
...