Рибонуклеозиды (на примере инозина)Статьи / Полупрепаративный микробиологический синтез биологически активных соединений, меченных стабильными изотопами водорода и углерода / Рибонуклеозиды (на примере инозина)
2
Н-меченый инозин.
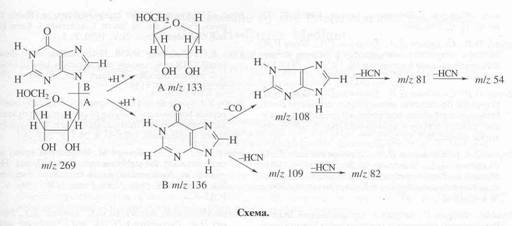
Уровень дейтерированности определяли методом масс-спектрометрии FAB. Фрагментация молекулы инозина при бомбардировании быстрыми электронами заключается в формировании ионизированного пика его молекулярного иона [М + Н]+ при m/z 269, который впоследствии распадается на фрагмент рибозы А при m/z 133 и гипоксантиновый фрагмент В при m/z 136, расщепляющийся в свою очередь на ряд менее низкомолекулярных осколочных фрагментов при m/z 54, ., 109 за счет элиминирования НСN и СО из гипоксантина (схема). Уровень дейтерированности инозина составил пять атомов из восьми детектируемых по скелету молекулы или 62.5% относительно общего количества атомов водорода в углеродном скелете молекулы. Примеси молекул с включением четырех (m/z при 273, 23%), шести (m/z при 275, 32%) и семи атомов дейтерия (m/z при 276, 20%) также фиксировались в масс-спектре (рис. 19). Возможные места локализации дейтерия определяли по тяжелым пикам фрагментов рибозы C5H9O4+ при m/z 136, 46% (вместо m/z 133, 42%) и гипоксантина C5H4ON4+ при m/z 138, 54% (вместо m/z 136, 50%) в масс-спектре. Исходя из полученных данных можно предположить, что три атома дейтерия локализуются в рибозной части молекулы и два - в гипоксантиновом остатке. Достоверность полученного результата подтверждалась присутствием тяжелых пиков низкомолекулярных фрагментов, продуктов распада гипоксантина при m/z 84, 42% (вместо m/z при 82, 40%), и 111, 49% (вместо m/z при 109, 45%).
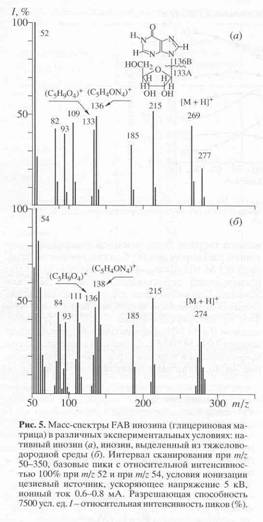
При анализе уровня дейтерированности 2Н-меченого инозина руководствовались следующими аспектами. Во-первых, изотопный полиморфизм свидетельствует о низкой специфичности изотопного включения дейтерия в молекулу инозина и, по-видимому, определяется условиями самого биосинтеза. Во-вторых, вследствие того что протоны (дейтероны) в С’1-С’5 положениях рибозной части могли происходить из глюкозы, предпологалось, что биосинтетическое включение дейтерия в рибозную часть молекулы инозина определяется в основном функционированием ряда процессов ГМФ-шунта, связанных непосредственно с ассимиляцией глюкозы. Вследствие того что глюкоза использовалась в протонированном виде, ее вклад в уровень дейтерированности рибозного фрагмента пренебрегался. Однако, вопреки этому предположению наблюдалось включение дейтерия в рибозный фрагмент молекулы инозина, что могло быть следствием сохранения доли минорных путей биосинтеза глюкозы из предшественников de novo. В-третьих, возможные (Н - 2Н) обменные процессы и изомеризационные превращения, происходящие во внутриклеточной среде в присутствии 2Н2O могли приводить к специфическому включению дейтерия по определенным позициям в молекуле инозина. Такими доступными позициями в молекуле инозина признаны прежде всего протоны при гидроксильных группах -ОН в рибозном остатке и имидазольные протоны при гетероатомах -NH (последние могут обмениваться на дейтерий в 2Н2O за счет кето-енольной таутомерии). Атомы дейтерия в остатке гипоксантина включались из 2Н-меченых аминокислот.
Смотрите также
Специальные варианты высокоэффективной жидкостной хроматографии
...
Тепловой эффект химической реакции
Тепловые
эффекты химических реакций необходимы для многих технических расчетов. Они
находят обширное применение во многих отраслях промышленности, а также в
военных разработках.
Целью
д ...
Химический элемент ванадий
В начале XIX в. в
Швеции были найдены новые богатые месторождения железной руды. Одна за другой
сооружались доменные печи. Но что примечательно: при одинаковых условиях
некоторые из ни ...